
I am grateful to Manhattan Community College, CUNY, for the 2009 Fellowship Leave that allowed me to complete this project.
Darwin documented his experiences on the Galápagos Islands from 15 September to 30 October 1835 in 40 primary texts and in nearly 200 references in his correspondence and private discourse. Therefore, the study of this crucial six-week period should not be limited to Chapter XIX, “Galápagos Archipelago,” of the 1845 edition of the Journal of Researches, popularly known as the Voyage of the Beagle. The notion that Darwin wrote briefly about the Galápagos experience in a single context has been inadvertently propagated on all levels of education. Popular high school and undergraduate biology texts, because of curricular requirements, tend to restrict their discussions to the Voyage of the Beagle, leaving it up to instructors to explain how ideas in evolutionary biology and geology stemming from the Galápagos period developed in later contexts (Koob & Boggs, 366-69; Miller & Levine 371-72, 406-09; Campbell & Reece, et al., 17-18, 23, 455-60, 487-92). This abridgement is also true for many authoritative studies, both general and specific (e.g., Mayr, 265, 408, 448; Ghiselin 25, 33, 34, 37; Abzhanov 1001-1007); however, the scholarship of Sulloway (1982-2009), of Grant and Estes (2000, 2009), and of others, in emphasizing that Darwin’s evolutionary thought developed gradually over the post-voyage decades, has extended the range of study to include both early and later texts, including notes and letters. This essay is intended as a guide to Darwin’s reflections on the Galápagos experience, from 1832 to 1876.
Figure 1. The Galápagos Archipelago, 600 miles west of Ecuador (Courtesy of smallshipsafaris.com). [Click on image to enlarge it.]
1. The Field Notebooks & Beagle Diary, autumn, 1835
Darwin filled fourteen notebooks on his shore excursions (Sept. 15-Oct. 30, 1835), while on the voyage of H.M.S. Beagle, 1832-1836. As he explored the islands, he recorded impressive sights in a series of pocket books, eighteen of which are concerned with observations on geology and natural history (R. D. Keynes, ed., BD. xiv). A notebook, titled “Galápagos, Otaheite Lima” (Galápagos Notebooks), the first document in the corpus, contains 33 pages on the excursion. Of the 33 pages, 27 in large part describe geology, particularly lava beds, craters, volcanic structures and gullies, and include measurements of rock-formation, altitudes and diameters, and classifications of mineral deposits. Though these Galápagos entries are fragmentary, they have descriptive value.
Unlike the field notebooks, the Beagle Diary was written in relative comfort on board or onshore, thus providing Darwin with the opportunity to organize field-work observations and to draw connections between disparate phenomena (Keynes, BD, xiv). As in the pedestrian notes, geology once again predominates, and Darwin would eventually devote a large volume to it alone: the 1844 Geological observations on the volcanic islands visited during the voyage of H.M.S. Beagle. Of the twenty-five chronological entries in the Diary’s Galápagos pages (Sept. 15-October 19), thirteen are concerned with geology and hydrography, three with ornithology, seven with botany, five with meteorology, and four with herpetology. In the Diary, zoology and geography were equally represented.
In the entries of 26-27 September and of 1 October where the phrase “centers of creation” appears, Darwin suggests that, contrary to popular thought, the islands were teeming with diverse forms of life. In many of these passages, he elaborated on the field notes; for example, he conjectured not only that volcanic structures had arisen from the sub-aqueous depths (BD, p. 356), but that a dynamic biological process affected living things (BD, p. 356). To understand this operative force, he compared the ecology of the island group to that of the Ecuadorian mainland. Four months later, he would advert to the central problem of whether Galápagos flora originated in South America or were peculiar to the Archipelago (CDC. 1, p. 485). The Galápagos commentary in the 1839 Journal and Remarks and in the 1845 Journal of Researches evidence Darwin’s consistent effort to understand the bio-geographical material he had amassed.
2. The 1839 Journal and Remarks
The 1839 Journal is the third in a three-volume set describing consecutive Beagle voyages: that of 1826-1830 (volume I by Captain Philip Park King et al.) and that of 1831-1836 (volumes II and III, by Captain Robert Fitz Roy and by Charles Darwin, respectively). The treatment of the Archipelago excursion in the 1839 Journal differs significantly, in form and content, from what Darwin had written about Galápagos in the Field Notebooks and in the Beagle Diary. The Journal was published before Darwin could fully appreciate what the huge collection of specimens had to communicate, a delay attributed to numerous misconceptions he had about island life-forms, notably in regard to tortoises and finches (Sulloway [1984]: 29). Aware of the organizational and interpretative complexity of the project, he began a fruitful collaboration with other naturalists (pp. 466-473; 8-18 October 1835), and, with their guidance, studied the compiled evidence systematically. In 1839, extended commentaries replaced the chronological ordering of events in earlier texts (only six dates, for example, are recorded in the Galápagos entry of 1839).
More than a repository of observed data and conjecture, 1839 furnished Darwin with the opportunity to study Galápagos biology. Of 40 passages of varying lengths, approximately 8% involves geology while 50% is given to zoology, a significant difference from how topics were apportioned in the field-notes and diary entries. At least twelve passages in 1839 bear directly on evolutionary biology. From Field Notebook, to Diary, to Journal, his intellectual approach gradually changed, the collector and observer becoming the hypothetically-deductive thinker. While describing the mundane topography of Chatham Island, Darwin had a profound insight, one that would inform his thinking for decades to come: “The natural history of this archipelago . . . seems to be a little world within itself; the greater number of its inhabitants, both vegetable and animal, being found nowhere else” (JR [1839], pp. 454-55).
The observant hiker, by 1839, assumed the dual role of superintendent and contributor to a grand project in natural history. Inklings of this great undertaking are detectable as early as 8 October 1835, in his “general observations” on the islands’ natural history. With less than two months to work on the islands, he had labored to amass “a perfect collection in every branch as time permitted”—a virtual impossibility, of course. So he would turn to shipmates and to colleagues in England. The floral specimens were given to the botanists J. S. Henslow and Richard Dalton Hooker; Henslow, for example, perceived that certain “weedy” plants found at higher elevations were probably new species, and that certain floral species, though in certain respects analogous to indigenous South American types, were nevertheless unique to the Archipelago (JR [1839], p. 460). To the perception of a life-force or natural law at work on the islands, Darwin added another mysterious characteristic of island life: organic beings were, at once, similar to mainland species yet obviously different from them.
The ornithologist John Gould, a renowned naturalist whom Darwin had enlisted in the project, tried to account for relationships, if any, between Continental and island species. On the principal islands, they had identified 26 discrete species of land-birds, of which 25 were hitherto unknown and presumably exclusive to the Archipelago (JR [1839], p. 461). In 1839 Darwin hypothesized that unique variations could be found in allied species throughout the Archipelago. He focused, tentatively, on the famous instance of the finches, the centerpiece of modern biological education, as a way of solving the problem. All of the finches he had collected agreed in beak morphology, in tail length, in general form, and in plumage. On the basis of his collection and on Gould’s advice, he noted important differences, such as females being gray or brown and mature males jet-black. All finch species, with two exceptions, fed in flocks on the ground and had similar habits. But subtle differences in beaks between particular species appeared to be the key to understanding evolutionary processes: “It is very remarkable,” writes Darwin, “that a nearly perfect gradation of structure in the one group can be traced in the form of the beak, from one exceeding in dimensions that of the largest gros-beak, to another differing but little from that of the warbler” (JR [1839], p. 461). Whether the comparative evidence could reveal a natural law or process, only intellectual labor would show. But, through observation and the comparative method, Darwin made considerable progress towards a synthetic proposition. In 1839, he had a working hypothesis (species evolved and both genus and species exhibited diversity); was able to compare living forms to each other physiologically; and had deduced that Galápagos life-forms resembled mainland forms yet, from island to island, were apparently unique to each ecological niche. The unanswered question before him was why animals and plants of the same genus living on different islands differed from one another.
Ideas on variation and migration described above were not restricted to ornithology. To uncover a general law or process of nature affecting all living things, Darwin had to consider these phenomena as they were manifested in other zoological Orders on the islands, such as the reptile, the “striking features” and great numbers of which he found quite impressive. He made the most of the opportunity to describe the feeding and reproductive habits of tortoises and of lizards, both terrestrial and marine, and to investigate the possibility that the laws governing the variation and distribution of reptiles were analogous, if not identical to, those affecting birds (JR [1839], pp. 462-73).
Darwin found that these premises applied to most living things on the Archipelago. With the exception of wanderers, a number of the animals on Galápagos were anomalies: on the one hand, appearing “peculiar to it,” but, on the other hand, “impressed by” South American forms. He found this to be true in the correspondence between island birds and those gathered while in Chile. “This similarity in type between distant islands and continents,” as well as between distinct species, was a pivotal observation. To reconcile the ideas of nature’s constancy and proliferative variability, he states “that the creative power has acted according to the same law over a wide area” (JR [1839], p. 474). Recapitulating the 1835 notion that there were in nature discernible loci of organic activity, he implies that an inherent natural law, imperceptible to the casual observer, was constantly at work, and that its effects were variegated.
An important premise of this Guide is that Darwin’s ideas on evolution did not come in a flash of insight but, rather, developed over time and not along a linear trajectory. Frank J. Sulloway propounds this idea as he clarifies our understanding of the finches’ place in the development of Darwin’s evolutionary theory, proving that information collected from the islands was deciphered gradually over decades. Contrary to popularly-held beliefs, the finches did not inspire Darwin’s earliest views of evolution, even after he was fully committed to the theory from 1837 on, and even though he was continually interested in ornithology. Darwin’s evolutionary views, as Sulloway argues, allowed for the understanding of significant finch-diversity on the islands, but gradually and retrospectively. In the Notebooks of 1837-1844, finches are mentioned but not explicitly connected with Galápagos. Sulloway points out that, in Darwin’s commentary on the transmutation of species—and on topics such as speciation, geographical isolation, and adaptive radiation--the finch collection, at the center of modern pedagogy, did not figure at all. In 1839, aside from a comment on the subgenera Geospiza, Darwin (according to Sulloway) had little else to say about finches.
In 1839, Darwin could say with certainty that the thirteen species of ground-finches, specimens assembled and profiled, presented “a nearly perfect gradation,” traceable from an extraordinarily thick beak to a very fine one. Compositely, these observations and contrasts strongly suggested that “certain members of the [ground-finch] series are confined to different islands” (JR [1839], p. 475). To prove this, Darwin admitted, a naturalist would have to collect specimens from geographically-diverse localities, an impractical undertaking. Nevertheless, he hypothesized (correctly) on the basis of the evidence at hand: if several islands were populated by species peculiar to each one but of the same genera, upon further inspection, “a wide range of character” in these insular species probably would exist; that is, nature seemed infinitely inventive, as far as could be discerned. How this was possible was yet unclear. Darwin reflects that this “curious subject” was too complex to be treated in the limited context of the 1839 Journal (JR [1839], p. 475).
3. Notebooks, 1837-1844; the Zoology Project, 1838-1844; & The Origin of Species Texts, 1842 & 1844
Prior to the publication of 1839, five books on the Beagle voyage, dealing specifically with Darwin’s collections, had been published. In 1839, six more would appear; and, before the 1845 redaction of the Journal, nine volumes more. While this monumental project was ongoing, Darwin contemplated the biological ideas and contradictions emerging from his original explorations. These ideas would be worked on in the private discourse, in a series of notebooks, and in the correspondence, from 1832 to 1845. Darwin’s notebooks (1837 to 1844) and drafts of The Origin of Species (1842 & 1844) form a bridge to the 1845 edition of the Journal of Researches. Below is a topical list of thirty-five Galápagos-related entries from nine notebooks:
-arbitrary nature of species: MAC167, QE109 -birds: B7, 55, 70, 98, 100, 103: C145, 209 -geographical dispersal: A46; B98, 100, 220; D65 -Galápagos plants: B157, 173; C184, 194; E100, 104; QE [15v} -South American character of species: B11, 98,103, 160; C54, 209; E100, 170 -species peculiar to separate islets: B7; D53; E32, 100; ZEd8 [Notebooks, 1836-1844]
One particularly incisive entry, MAC167 (late 1838), is a marginal abstract reflecting on the natural theologian John Macculloch’s Proofs of the Attributes of God from the Facts and Laws of the Physical Universe, being the Foundation of Natural and Revealed Religion (1837). Darwin’s commentary in MAC167 is clearly an extension and landmark of the work up to 1839:[We] can deduce why small islands should possess many peculiar species. For as long as physical change is in progress or is, present with . . . respect to new arriv[als], the small body of species would far more easily be changed.—Hence the Galápagos [Islands] are explained. On distinct Creation, how anomalous, that the smallest newest, & most wretched [island] should possess species to themselves.—Probably no case in [the] world like Galápagos. No hurricanes.—[islands] were never joined, nature & climate very different, from adjoining coast. Admirable explanation is thus offered. [MAC167, CDN., p. 640]
Darwin proposed, in MAC167, that small islands were particularly conducive to the engendering and transmutation of species. He isolated three ideas that could account for the dynamic speciation on the Archipelago: (1) if new creatures were to arrive when ecological change was in progress, and if they were to commingle with indigenous species, the likelihood was that new species would be generated; (2) in the Galápagos group, the impulse toward variation was most intense on a relatively recent geological formation (e.g., a submerged islet, volcanically thrust upward), and especially on one having never been connected to South America; and (3), in any discussion of species, the nature and climate of the main islands, which varied considerably between each other, had to be taken into account; nor could oceanographic and climatic conditions explain the similarity between island and mainland types since conditions actually inhibited the transportation of mainland seeds and the migration of animals to the islands.
Darwin wrote two sketches preliminary to the 1859 Origin of Species, an initial draft in 1842 and a revised one in 1844. In 1844 he re-explored Galápagos traits and bio-geographical problems. One was the logical expectation that life-forms on small islands, situated relatively close to land, will have similar fauna, which would validate the possibility of past migration from the mainland. Superficial similarities notwithstanding, the islands, as already stated, have a large number of “entirely peculiar fauna” (1844 Essay, p. 159), which held true for birds, reptiles, shells, and fish. What accounted for the fact that, despite their uniqueness, fauna on the Galápagos also resembled American types? Darwin initially proposed, but would back away from, the idea that groups of islands, such as the Galápagos Archipelago, constituted “small but well-defined sub-divisions of the larger geographical divisions” (1844 Essay, p. 159). Could the Archipelago, then, be thought of as a cluster of component islands or, in totality, as a South American cluster of satellites? This hypothesis was not convincing, since the islands were volcanic and craterous and, unlike the mainland of the Ecuador, had to be of recent origin. For Darwin, since conditions favored neither the migration of animals and birds (650 miles west of Ecuador), nor the transportation of seeds, and if the islands were younger than the Continent itself, then its biological tenants had to have been newly formed and indigenous. Since so many species seemed to be “peculiar” to their respective environments, Darwin had reason to believe that they had been “produced on this very spot,” an extraordinary intuition (1844 Essay, pp. 159-60). The answer could lay in bio-geography: what accounted for the influence of the soil and climate on the islands, both of which were considered dissimilar from that of the Ecuadorian coast? In terms of general form, Galápagos species undoubtedly corresponded to “closely allied type[s]” on the mainland (1844 Essay, pp. 159-60). At each phase and retracing of Darwin’s thought, we find an inquiring mind attempting to incorporate disparate observations and elucidating glimpses into a coherent description of natural reality.
When Darwin arrived at cognitive impasses, he sought alternative routes. One such re-routing is obtained through analogy, the inference that if two things are similar in certain ways, they must also be similar in others. Thus, Darwin compared Galápagos and Ecuador to other archipelagos and corresponding mainlands. The juxtaposition of Galápagos-Ecuador to Cape de Verde-Senegal, for example, was geologically reasonable, as the physical conditions of Galápagos closely resembled those of the Cape de Verde volcanic group. Another encouraging similarity was that the flora and fauna on each island in the Cape de Verde group (350 miles West of Dakar, Senegal, in the Atlantic Ocean) bore traits that were adapted to that particular environment, an observation that held true for the main islands of the Galápagos group. Darwin found further evidence of ecologically-adapted characteristics in the fauna of the sub-tropical Canary Islands off the coast of Morocco in the Atlantic Ocean. Animals on these islands were not only allied to Southern European types, just as the inhabitants of the Cape de Verde Islands were allied to African species, but they also had traits unique to their respective island habitats. The analogy raises the question: how do bird species on the three Archipelagos compare to each other? Darwin was able to make broad linkages. Three factors were common to the flora and fauna of these island groups: all “organic beings” (to use Darwin’s serviceable phrase) were similar to correlatives on the respective mainland; unique forms inhabited the islands, having no counterpart on the mainland; and each island exhibited life-forms with traits characteristic of, or adapted to, it in particular. For reasons as-yet unknown to Darwin, geography had an undefined bearing on species.
Darwin and his colleagues, at this time, were set to work on the immense zoology project. The specimens collected on the voyage, as mentioned above, needed scientific cataloguing; and a catalogue, both texts and images, was to be published. It was obviously a monumental undertaking, far exceeding the ability of any one individual, even one as indefatigable as Darwin. Yet he still managed to superintend the collaborative project and to be a major contributor. Five other naturalists joined him. The entire work, written from February 1838 to October 1843, would consist of five parts and nineteen numbers or subsidiary texts (see Texts and Correspondence). Darwin was concurrently at work on a full-scale study, unofficially the sixth part of the project, entitled Geological observations on the volcanic islands visited during the voyage of H.M.S. Beagle, which would be published in 1844; an entire chapter was fittingly allotted to the geology of the Archipelago, particularly to Chatham, to Albemarle, and to James Islands (recurring to the Field Notebooks) (pp. 97-116), and it would be interesting to trace its genesis from 1835 to 1844, that is, from the Field Notebooks to Geological Observations. Concurrently the project editor, superintendent, and contributor, he also found time to write a geological introduction to Part I, the Fossil Mammalia (pp. 3-12), a geographical introduction to Part II, the Mammalia (pp. i-iv), as well as remarks on habits and migratory ranges throughout the Mammalia and the Birds, along with notes in the Fish and the Reptiles (Freeman [1977], as cited in The Complete Work of Charles Darwin [2009]).
Five eminent collaborators participated in the Project: the comparative anatomist Richard Owen (1804-92) (Fossil Mammalia); the naturalist and founder of the Entomological Society, 1833, George Robert Waterhouse (1810-88) (Mammalia); the self-taught ornithologist and artist John Gould (1804-81) (Birds); the naturalist and clergyman Leonard Jenyns (1800-93) (Fish); and the professor of zoology Thomas Bell (1792-1880) (Reptiles); in addition, John Stevens Henslow (1796-1861) and botanist Joseph Dalton Hooker (1817-1911) were responsible for plant specimens (Biographical Register and Index to Correspondents, CDC. 1, pp 611-61). Their publications appeared mainly in periodicals. Henslow, who had extended and remodeled the Cambridge Botanic Garden, wrote about native plants on Keeling Island (“Forula Keelingensis: An account of the native plants of the Keeling Islands” [Annals of Natural History, [1838] 1:337-47]); and Hooker, whose interest was plant geography and classification, contributed. “An enumeration of the plants of the Galápagos Archipelago, with descriptions of those which are new” (Transactions of the Linnaean Society of London [Botany], [1847] 20:163-233); and “On the vegetation of the Galápagos Archipelago as compared with that of some other tropical islands and of the continent of America” (Transactions of the Linnaean Society of London [Botany], [1847] 20:235-62) (Bibliography, CDN, pp. 653-92). The aesthetic and informative value of these texts is unquestionable. Gould, for example, thoroughly classified and described the Genus Geospiza, a finch indigenous to the Archipelago and contributed an important gloss on the biological commentary in the 1845 Journal of Researches. With this information systematically ordered, Darwin was able to maintain his focus on unanswered questions pertaining to variation, migration, distribution, habitat, and species.
The voluminous Zoology Project was completed in 1843, the Geological observations appeared in 1844, and Darwin was preoccupied with the 1842 and 1844 drafts of The Origin; in addition, he labored on a redacted version of the 1845 Journal Researches (previously the Journal and Remarks). At this juncture, and with observations and specimens in relative order, his critical enquiry resumed with even greater clarity.
In the 1845 Journal of Researches, the sixth major installment of the corpus (if the Zoology and Geological observations are grouped together), Darwin reiterated the threefold puzzle of the Galápagos islands: (1) many life-forms were found there and nowhere else in the world; (2) closely-related inhabitants of the principal islands varied subtly from one another; and (3) all known creatures were, in general respects, similar to mainland correlatives, even though the South American continent was 500 to 600 miles away. The Archipelago, undeniably, was a microcosm of remarkable organic creativity, perhaps even an oceanic satellite of America, from which “a few stray colonists are derived.” Wherever original migrants originated, and whenever they had arrived, remained mysteries. Though migrations had undoubtedly taken place, the idea of the Archipelago as a microcosm in which new life-forms were engendered was foremost in his mind (JR [1845], p. 377-78).
Statistical data, a new Journal feature, was now employed to support Darwin’s evaluations in 1845 chapter (Chapter XVII, pp. 372-401). Surveying the species and habitat data, organized by the zoologists and botanists from 1838 to 1843, he targeted the question of why the islands’ inhabitants were biologically unique, but now his empirical observations could be mathematically tested. To present numerical evidence and interrelationships, he employed two metrics: proportionality, the relation of one part to another or to the whole with respect to magnitude, quantity, and degree; and tabulation, the computation and presentation of data in lists and column, arranged for easy understanding. Reproduced below and redacted from the 1845 Journal, this Table demonstrates the law of species distribution in regard to island flora gathered over a few days. For the first time in a publication, Darwin was able to quantify his empirical and analogical observations:
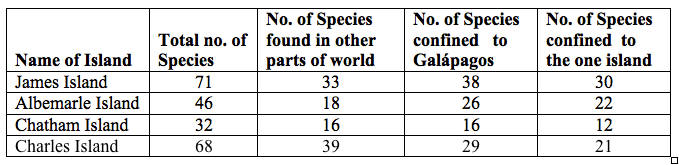
Figure 2.Distribution of Flora (Modified version)
Here is what Darwin and his colleagues established. Approximately one-half of the total plant species sampled on the four principal islands (first column) were unique to the Archipelago. Fifty-four percent of the James Island species (38/71) were indigenous to the Galápagos Islands as a whole and not found anywhere else. That nearly 80% of the plant specimens (30/38) gathered on James Island was found only on that island, supported the hypothesis that each island was a unique ecosystem. The same proportions were true for Albemarle Island, the largest in the group: whereas 56% of the floral species there were unique to the Galápagos, once again, nearly 80% of the plant species were exclusive to Albemarle. The statistics are astonishingly consistent for Chatham Island as well: 50% of the flora gathered there was typical only of Galápagos, but approximately 80% of these variant specimens lived only on Chatham. Charles Island fits the speciation pattern as well: 42% of its retrieved plant species were unique to Galápagos, while about 70% inhabited only Charles Island (along with Hood, Charles was southernmost in the group and among the smallest).
To account for the consistent factors, regarding the distribution and the exclusivity of flora and of fauna, Darwin considered returning to the oceanographic argument. The transportation of insects, seeds, and animals depended on weather, ocean and wind currents, and on human visitors. Because strong currents ran in westerly and W.N.W. directions, they likely separated southern from northern districts of the Archipelago. Accordingly, the islets Culpepper, Weman, Abingdon, Bindloe, and Tower, in relation to the main islands could be considered a distinct faunal and floral district. This supposition seemed reasonable. But what accounted for the marked, specific variations between closely situated, southerly islands, such as James and Albemarle? The lack of cross-fertilization between the two islands could be explained by the strong northwest ocean current, effectually separating them. Meteorology, however, did not support the migration theory. Since gale winds (30-60 mph) were not typical of these islands, birds, insects, and light seeds could not have been blown between them. Additionally, the great ocean depth between the islands and their apparently recent geological origin rendered it unlikely that they were ever united as a single landmass, thus precluding the possibility of terrestrial migration from one to the other. Since an explanation of how organic life on island and Continent related to one another remained elusive, Darwin found himself at another momentary impasse.
Darwin re-directed his approach to the problem of the uniqueness of life-forms on each island and their unmistakable similarities to South American types. His rhetorical maneuver, at this juncture, was to re-define the Archipelago, ecologically, not as an aggregative satellite of South America, but as an array of biogeographically-distinct satellites; thus, he viewed the nine islands, 50 islets and reefs, scattered over an area of nearly 200 miles or 320 kilometers, as individual entities and as distinct ecological systems (JR [1845], pp. 396-98).
In The Origin of Species ([1859], pp. 398-402), as in earlier drafts, Darwin outlined what he had been inferring since 1835. He compares the Galápagos-Ecuador (the coast of Quito, Ecuador, is about 750 miles or 1200 kilometers due East of Albemarle), to other island groups and their respective mainland. The zoology and flora of the Cape de Verde Archipelago related to coastal Senegalese species, Darwin points out, in a manner similar to that in which Galápagos life related to corresponding life-forms on the Ecuadorian coast. But analogy alone could not reveal why life-forms were unique to immediate environments yet, at the same time, similar to related species on nearby islands and on the mainland. The Cape de Verde–Galápagos analogue, however, was a familiar starting point: first, both were volcanic, had similar climates, soil, altitudes, and areas; second, plants and animals in the Galápagos resembled types in Ecuador, which held true for Cape de Verde and Senegal. That much, the analogy could reveal. But neither geography nor “the ordinary view of independent creation,” offered Darwin a satisfactory explanation as to why Galápagos life was so unusual.
The problem with Darwin’s reasoning, up to 1859, was that he ascribed to the physical condition of a country more significance in the formation of species and of variations than was warranted (OS [1859], pp. 398-402). The real power at the center of creation, he discovered, was not inherent in the environment but, as modern historians have shown, was generated by the life-and-death struggle between competing species and Orders and ultimately was a matter of the inheritance of traits conducive to survival. This was a revelation to Darwin, but one that manifested itself through accretion and critical reasoning: survival was determined through successful competition.
Competition between species on distinct islands, Darwin surmised, seemed to account for indigenous modifications, even if an original seed or animal derived from a common source on the mainland. Darwin understood the power of competitiveness in the late 1830s when he referred to “the wars of organic being” (Notebook, C73, p. 262) and used the simile of “a hundred thousand wedges” to describe the unrelenting pressure to adapt, to propagate, or to die out (D134-135, pp. 374-376). Dissimilarity, he learned, was a manifestation of adaptability: if a South American seed germinated on one island and if it subsequently flourished, it had become adapted to environmental factors and to its teeming, predatory habitat. The windblown seed landing, let us say, on Albemarle soil, would be exposed to conditions in a habitat unique to Albemarle. A newcomer, under each new set of conditions, would then undergo the challenges of the respective biosphere. Those that survived produced similarly or better adapted progeny, and so the process continued, with extinction an ever-present possibility should an ecological imbalance subvert the adaptive balance.
Natural Selection, or the Survival of the Fittest, ultimately furnished the key to understanding adaptation and variability (OS [1859], pp. 80-131). Involving the preservation of favorable individual differences and variations and the destruction of injurious ones, it did not induce variability itself, as would an active, external force, but rather implied only the preservation of variations that, in certain respects, were “beneficial to the being under its conditions of life.” All living forms, regardless of location, were subject to these conditions. Climate was not necessarily the most important factor in the struggle for life. Darwin explained that, “when a plant or an animal is placed in a new country amongst new competitors, the conditions of its life will generally be changed in an essential manner, although the climate may be exactly the same as in its former home.” Invariably, if one species has an advantage over another, it will in a brief time supplant its rivals. An intriguing question for classroom use would be as follows: if one were to transport a particular bird species from one island to another in the Galápagos chain, what would determine its ability to survive and prosper? If it died out amidst birds of the same genus, suggest reasons why the migrant bird not succeed?
From 1860 to 1878, Darwin returned, intermittently, to lessons learned on the Galápagos Islands. He did so in five major books and in more than two-dozen letters and related contexts, all of which are listed below.
Related Material
- Primary Darwinian texts for Darwin and the Galápagos
- Secondary Texts for Darwin and the Galápagos
- A Chronological List of Primary Darwinian Texts & Related Correspondence
Last modified 23 August 2013